
Every breeder colony sould bee VSH tested
from Erik Österlund
VSH-trait
- VSH (Varroa Sensitive Hygiene) is a trait in worker bees.
- Worker pupae with Varroa mites that have offspring are cleaned out.
- The fertile female mite survives, but all offspring dies, as well as the worker pupa.
- Worker pupae with Varroa mites that don’t have offspring are not cleaned out.
- VSH-trait is governed by two genes. As the genes comes in double (in two sets of chromosomes) there are two pair of genes, which makes four alleles (variants of genes).
- The alleles that contribute to VSH are additive, not recessive or dominant. Two is better than one, etc.
- Two alleles in a bee makes it show the VSH trait in such a strong way the colony acts as resistant
The above list is some of the things I learned from John Harbo when he was lecturing in Sweden recently. That VSH trait is governed by two pairs of genes is a working hypothesis that is contributed by breeding tests. No observation contradict this.

This makes the VSH-trait to something different compared to the hygienic trait keeping bee colonies free from American foul brood and chalk brood. This later hygienic trait is most often tested checking how fast bees clean out freeze killed brood (or pin killed). The working hypothesis is here that the trait is goverend by three gene pairs and this hygienic trait is contributed by recessive alleles (gene variants), which makes it necessary that individual worker bees have all six alleles to show the trait. I have understood that the more individual bees in a bee colony that have all six alleles the more effective is the colony in this hygienic trait.
But with VSH you can for example have a queen with two alleles that are mated to drones with two alleles. Thus the virgins from such a colony always will have two alleles. Those virgins can mate to any kind of drone, with or without VSH-alleles, and still show the VSH-trait – AND be Varroa resistant.
On the other hand it is not necessary that all individual bees in a bee colony have two VSH-alleles for the whole colony to be Varroa resistant. It is unclear how many are needed. But in a test where a resistant colony and a non-resistant were combined – the combined colony was resistant. This means that you don’t have to use an inbred breeder colony with all four alleles to produce varroa resistant bee colonies. If your breeder colonies have a high VSH-value and you open mate the daughters in an area dominated by drones from colonies with high VSH-index it’s good chance the queens produced will give Varroa resistant colonies, if not reinvaded heavily by mites…
You can test your bee colonies for the VSH-trait and breed from the best and with a good strategy develop the Varroa resistance in your bee stock, even if you don’t use instrumental insemination. But if you do, you can develop a strain quicker with higher number of alleles.
All bee stocks have the VSH trait, at least at a low degree. The alleles are there, but spread out in the population. Not concentrated enough in the bees in a non-selected stock. Also mites get old and stop reproducing. So “background noise” of non reproductive (infertile) mites in a colony is about 10%. Maybe even up to 25%. If an individual bee have one VSH allele the VSH trait doesn’t show up in that bee. I think that’s the info I got.

After retirement John Harbo bought queens from Glenn Apiaries in California to start his own bee business. He continued breeding VSH bees and he doesn’t treat his bees against the Varroa mite. He doesn’t need to. He is now running his bee business together with his wife Carol.

Harbo explained a simple test every beekeeper can and every bee breeder should do. He uses a microscope with 0.5 magnification of the eyepiece. Thus getting maybe about a total of 5x magnification. The working distance under the microscope need to be about 10 centimeters (4 inches).

I use a reading magnification lense for elderly people with a ring lamp (much cheaper), plus reading glasses (2.5x).

(Click on the picture to enlarge it and on back arrow up to the left to come back to the text.) When the pupae are at least 4 days old and the eyes are getting dark is the best time to check them. With tweezers around the head (or somewhat further down) you pull out the pupa, look at the pupa for mites and mite feces (mites that don’t reproduce often put there feces on the pupa instead of 10 or 2 o’clock closer to the bottom of the cell but on the cell wall. You then look in the cell for mites and mite feces. Often you see white remnants of the pupa on the bottom of the cell. The mite feces are somewhat whiter than pupa remnants and more up on the cell wall, but still close to the bottom.
What I saw when I did my first test was that mites with good reproduction and whole varroa families, those mites were very active and ran around on the bottom. Mites with just one type or very few offspring moved slowly in comparison. Which also did the mother mites with no offspring, that didn’t reproduce. The mite feces amount in such cells with no reproduction seemed to be smaller.
- If a bee colony shows 100% VSH trait – 100% of the pupae with mites (of the total number of pupae with mites, that is all of the pupa with mites), the mites in those do not reproduce, they have no offspring – the bee colony have all four alleles for VSH.
- If a bee colony shows 60% VSH-trait (60% of the pupae with mites have no offspring), it has three alleles.
- If it shows 30% VSH-trait, it has two alleles – and is varroa resistant.
- If a colony shows 25% and below it has no breeding value according to Harbo. The figure is too close to ”background noise” of infertile mites of other reasons than influence of VSH-trait.
If 25% would be the best I could come up with I would personally use it anyway as a breeder if it is a good colony in other respects. I would also consider the possibilitty to increase other efforts of enchancing the Varroa resistance in my stock.
Why did I test the colony I did test as the first one of mine? Because it showed very little signs of varroa present. It is very good in other traits. It didn’t need Varroa treatment when neighboring colonies did. It showed 8 pupae with mites out of 110. 2 had whole Varroa families, 1 had just a male offspring. 1 had only two female daughters. The rest no offspring. This makes 50% VSH. Time will tell how resistant the colony is. It’s on small cells as well.
The VSH test is so easy and so valuable, that every bee breeder should tell the customers what VSH value their breeders have. And every queen customer should ask for it.
------
Responses to Varroa by honey bees - John Harbo
DOI 10.3896/IBRA.1.48.3.02
ORIGINAL RESEARCH ARTICLE
Responses to Varroa by honey bees with different levels of Varroa Sensitive Hygiene
John R. Harbo1 and Jeffrey W. Harris1*
1USDA/ARS Honey Bee Breeding, Genetics and Physiology Lab., Baton Rouge, Louisiana 70820 USA.
Received 6 January 2008, accepted subject to revision 14 May 2008, accepted for publication 17 August 2008.
*Corresponding author: Jeffrey.Harris
Summary
Mite resistance that we had earlier called suppression of mite reproduction (SMR) is a form of hygienic behaviour that we have named Varroa Sensitive Hygiene (VSH). With VSH, adult worker bees (Apis mellifera) disrupt the reproduction of parasitic mites (Varroa destructor) by removing mite infested bee pupae from their cells. This study determines which brood cells are targeted by bees with VSH and which are not, and describes the relationship between brood removal and the sudden prevalence of sterile mites. We produced 26 colonies with different levels of VSH by backcrossing 14 queens from a high line, H (100% expression of VSH), and 12 queens from a low line, L (no VSH), to drones produced by an HL (high × low) queen. Because each of the 26 queens was mated to one drone, the resulting colonies were expected to represent the complete range of variability (0 to 100% of the alleles for VSH). To estimate brood removal, we measured mite populations in capped worker brood that was 0 - 3 days postcapping and again 7 days later when the cohort was aged 7 - 10 days postcapping. We correlated removal of mite-infested brood with the relative presence (at 7 – 10 days postcapping) of three classes of foundress mites: (1) viable: those with at least one daughter that could mature before emergence of the host bee; (2) nonviable: those with progeny but with no daughters that could reach maturity; and (3) no eggs: those with dead or nonovipositing foundress mites. As the rate of removal increased, both classes 1 and 2 showed significant declines, whereas class 3 was unchanged. Therefore, oviposition of the mite or something associated with mite oviposition provides the stimulus for bees with the VSH trait to remove mite infested pupae.
Introduction
We selected honey bees for resistance to Varroa destructor Anderson and Trueman. Thanks to the use of single drone inseminations, we quickly found resistance (near elimination of the mite population). We called this characteristic Suppressed Mite Reproduction (SMR), because when we examined worker bee pupae that were 7-11 days postcapping, most or all of the mites had no viable progeny (Harbo and Harris, 1999). Studies have shown that this condition was caused by the removal of mite infested brood by adult bees (Ibrahim and Spivak, 2004, 2006; Harbo and Harris, 2005). This removal of mite infested pupae is apparently limited to worker pupae, and although bees may sometimes remove mite infested drone pupae, they do not significantly reduce the infestation in drone brood (Harris, 2008). Removal of mite infested pupae by adult bees explains why mite resistance was not observed until about six weeks after introducing a mite resistant queen, why the mites disappeared so suddenly, and how mite populations could recover in colonies where resistant queens were replaced by susceptible queens (Harbo and Harris, 1999; Harris and Harbo, 2000). We therefore renamed this mite resistant trait Varroa Sensitive Hygiene or VSH because it appears to be a form of hygienic behaviour. The term SMR, once useful in describing our breeding work, did not accurately describe the behaviour of the resistant bees.
Honey bees sometimes destroy and remove their brood in (what seems to us to be) an attempt to eliminate disease, parasites, or dead bees from their brood cells. The removal of bee pupae infested by V. destructor may be comparable to the removal of brood in cells infected with American foulbrood bacteria (Woodrow and Holst, 1942; Rothenbuhler, 1964). Rothenbuhler described this removal of diseased brood by adult bees as hygienic behaviour, and the term hygienic behaviour has since been expanded to include the removal of brood killed by freezing (Newton et al., 1975), brood killed by puncture (Spivak and Downey, 1998), brood infested with chalkbrood (Gilliam et al., 1983), brood invaded by the greater wax moth, Galleria mellonella (Corréa-Marques and De Jong, 1998; Villegas and Villa, 2006), or brood infested with the small hive beetle, Aethina tumida (Ellis et al., 2003, 2004; Neumann and Härtel, 2004). Thus, hygiene in bees has come to mean the detection and removal of diseased or dead brood by adult bees.
These different types of hygienic behaviour have similarities but also some differences. Spivak and Reuter (2001) found that colonies selected for rapid removal of freeze killed brood were more resistant to American Foulbrood (AFB). Studies that correlated high and low levels of hygiene between freeze killed brood and chalkbrood did not find close correlations (Taber, 1986; Spivak and Gilliam, 1993), but a group of colonies selected for freeze killed hygiene was overall more resistant to chalkbrood than was another group of colonies that had been selected for poor hygiene for freeze killed brood (Spivak and Gilliam, 1993; Spivak and Reuter, 2001). Similarly, VSH has some commonality with the other forms of hygiene but also some differences. For example, VSH produced intermediate phenotypes that could be best explained by additive alleles (Harbo and Harris, 2005a), whereas the hygienic behaviour responsible for resistance to American foulbrood (Rothenbuhler, 1964) was found to be a recessive trait. Hygienic removal of freeze killed brood seems to be controlled by the quantitative effects of multiple genes (Lapidge et al., 2002).
The purpose of this paper is to define the VSH trait as precisely as possible. VSH has two general elements: the removal of infested brood cells and a high frequency of nonreproducing mites. In early studies (Harbo and Hoopingarner, 1997) the prevalence of nonreproducing mites was strongly correlated with reduced mite populations, whilst freeze killed hygiene was not. The prevalence of nonreproducing mites was therefore used as the basis of our selection. We don’t know if this prevalence is a critical component of mite resistance, but it is a characteristic that defines VSH. A clear description of VSH should enable others: (1) to reproduce the trait;
(2) to describe variants or components of VSH; and (3) to recognize mite resistant traits that are partially or entirely independent ofVSH. Specific objectives of this study were: (1) to define which mite-infestations are targeted by VSH, and which are not; (2) to describe the relationship between brood removal and the relative presence of nonovipositing mites; and (3) to describe mite resistance qualities in colonies that have less than 100% of the alleles for VSH.
Materials and methods
We produced a group of 26 colonies that would represent all levels of expression of VSH.Fourteen colonies had queens produced from a line with 100% VSH expression (high line, H), and 12 had queens produced from a line that did not express VSH (low line, L). Each of the 26 queens was backcrossed to a single drone produced by an HL queen (a daughter of both lines H and L). Because spermatozoa from a haploid drone are all genetic replicates of the haploid egg from which the drone developed, the 26 drones represent 26 randomly segregated gametes from that queen. The drones should therefore range from having 0 to 100% of the alleles for VSH, and probability predicts equal numbers of drones with zero and 100 %. With this design, colonies in group H should have 50 – 100% of the alleles for VSH and those in group L should have 0 – 50%.
The test queens were introduced to colonies in July and early August. Colonies were evaluated for brood removal after the test queens had been laying in their colonies for at least six weeks. In this way all worker bees that are 0 – 3 weeks old would be daughters of the test queen.
Mite populations in newly capped brood
Measuring the population of mites in newly capped brood was our first step in estimating the removal of brood by bees with VSH. We sampled worker brood that was 0 - 3 days post-capping to estimate the baseline population of mites in each colony. We identified worker brood 0 - 3 days post-capping by examining only capped brood that had not yet shed their larval skins. Because most mites in the 0 - 3 day (initial infestation) group had not yet begun to lay eggs, this sample could only measure the rate of mite infestation. The same cohort of brood would be sampled again on the same comb seven days later to assess the mite population as well as the reproductive history of the remaining mites.
If fewer than 5% of the newly capped cells were infested, the colony was inoculated with further mites. We inoculated combs with mites by removing one comb of brood from a test colony and placing it into a highly infested colony for three days. The comb needed to have uncapped larvae that would be capped (and therefore infested with mites) during the next three days. We increased the effectiveness of a mite donating colony by caging the queen about two weeks before we gave combs to them from the test colonies. Caging was helpful because: (1) the donor colony would have no larvae to attract mites away from the target cells; (2) the donor colony would have a higher proportion of mites residing on adult bees and therefore available to invade; and (3) the caged queen would not contaminate the donor frame by laying eggs in it (in case we wanted to retest the colonies later). After three days, inoculated combs were removed from the mite donor colonies, the mite population in those combs was measured by examining 200 cells that were 0 - 3 days postcapping, and the combs were returned to their respective test colonies.
Measuring brood removal
Throughout this paper we refer to a measurement of brood removal. We measured a change in the frequency of mite infested brood and assumed that the disappearance of infested pupae was the removal of bee pupae by adult bees. If someone learns that the infested pupae disappeared by some other means, our analyses would still be valid but references to removal would need to be changed.
To measure brood removal, we sampled mite populations a second time when the cells were 7 – 10 days postcapping. We identified pupae of this age by their colour – purple eyed and white body to tan bodied pupae. We measured the infestation rate of the cells, the number of progeny, and the level of development of the progeny. We had to examine 428 ± 180 (mean ± SD) cells per colony in order to find an average of 20 ± 8 infested cells from each of the 26 colonies. A sample consisted of about 200 cells (more for the second sample, as explained above). Each sample counted the number of infested cells and the number of uninfested cells. We used the uninfested cells as our benchmark and produced an infestation term for each sample (number of infested cells per 100 uninfested cells).
Hypotheses tested
Our primary objective was to determine which, if any, of the mite infested cells were targeted by VSH. We addressed four possibilities:
(1) bees remove pupae at random; (2) bees remove all infested pupae equally; (3) bees only remove mite infested pupae that contain at least one viable daughter (one that could reach maturity before emergence of the host bee); or (4) bees remove all pupae with mites that lay eggs.
The first hypothesis was addressed by comparing mite removal in all colonies using Analysis of Variance (SAS 2000). Analysis compared the H and L colonies to determine if the removal of infested cells was different in the two groups and if each group was significantly different from zero. To adjust for non-normality of ratio data, each removal percentage was transformed to log10 of the removal rate + 1. Regression analyses were used to evaluate hypotheses 2 – 4. By evaluating survival of mites from each of three reproductive classes (mites with viable offspring, mites with nonviable offspring, and mites that lay no eggs), we could determine whether the survival of each class was affected by changes in the rate of removal of infested cells.
Regression analysis was also used to measure the correlation between the rate of removal of mite infested pupae and the prevalence of sterile and dead mites. We measured the frequency of sterile and dead mites (nonovipositing mites) by dividing the number of pupae with ovipositing mites by the number with nonovipositing mites. We called this measure the mite ovipositing ratio (OVR) and it can only be measured >7 days postcapping. A typical susceptible colony will have an OVR of 6 (6 pupae with reproducing mites for every infested pupa with no mite oviposition). Mite resistant colonies have an OVR between 0 and 1.5.
Response to Varroa by bees with VSH159
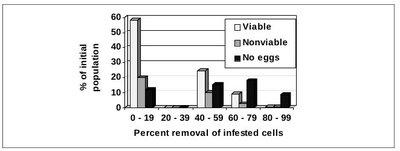
Fig. 1 Reproductive history of mites that survived hygienic removal. In each colony, every mite-infested cell that survived was sorted into one of three reproductive classes. With no mite removal, the sum of the 3 reproductive classes (the sum of the columns in each group) would equal 100%. The midpoint of the first group (0 – 19%) is 10% removal, so the sum of the columns is 90%. The midpoint of the last group (80 – 99%) is 90% removal, so the sum of the columns is 10% (90% of the infested cells in that group did not survive). This figure shows the relationship between the rate of removal of mite-infested brood and the mite reproduction in the brood that survived. Two classes of mite reproduction (viable progeny and nonviable progeny) decreased as the rate of removal increased, but there was no change in the survival of cells with mites that laid no eggs. Each set of columns represents the average of a group of colonies whose removal rates fell into that range. There were 8 colonies in the 0 – 19% range, none in the 20 – 39% range, and 5, 7, and 6 colonies in the higher ranges, respectively. Statistical analyses presented in the text used the exact removal rate for each colony rather than the grouped data.
Results
Bees with the VSH trait preferentially removed mite infested pupae when presented with infested capped worker brood. In particular, brood in the H colonies had a significant reduction in the number of infested cells (mean ± SE = 78 ± 7%) (df = 24, t =10.5, P <0.0001). Random removal of pupae would have resulted in no change in the infestation rate. Therefore, hypothesis 1 was rejected. Colonies in the L group also had an overall loss of infested cells (16 ± 7%), but it was not significantly different from zero (df = 24, t = 1.85, P = 0.08). The two group means were significantly different (df = 1, 24; F = 33.7; P < 0.0001).
Hypothesis 3 was rejected because the survival of the two classes of ovipositing mites significantly declined as the removal of mite infested cells increased (Fig.1). The relationship between brood removal and the survival of the viable class of mites was y = -0.62x + 56 where y = the percent survival of the viable class of mites and x = the percent removal of infested pupae (F = 196.0; df = 1, 24; P < 0.0001). The relationship was similar for the nonviable class of ovipositing mites, y = -0.21x + 20 (F = 42.6; df = 1, 24; P<0.0001).
Hypothesis 2 was rejected and hypothesis 4 was accepted because the no eggs class produced a slope that was not different from zero, y = -0.05x + 15 (F = 2.2; df = 1, 24; P < 0.15), and therefore showed no sign of removal. The constants (56, 20, and 15 %) in these equations estimate the relative presence of these three classes of mites in colonies that do not remove mite infested pupae (Fig. 1).
The relationship between brood removal and the mite ovipositing ratio
There was a strong linear relationship between percent removal of mite infested pupae (x) and the ovipositing ratio (y) (y = -0.057x + 5.8) (F = 27; df = 1, 24; P<0.0001; R2 = 0.52). Assuming that hypothesis 4 is correct and that 15% of the mites in all colonies would be nonovipositing if there was no brood removal, our predicted relationship would be y = -0.067x + 5.7. The constant or zero intercept is the mite-oviposition ratio (OVR) (85/15) when removal (x) = 0. When x = 85 (all ovipositing mites removed), the OVR should equal zero (0/any number >0). Therefore, the predicted slope (b) = -0.067. Both slope and intercept of the predicted line fall within the standard errors generated from the data (SE for slope and intercept are ± 0.011 and ± 0.7, respectively). Since the relationship between brood removal and the mite oviposition ratio are so close, we conclude that VSH can be estimated by measuring either of them.
Discussion
The removal of infested pupae and the nonremoval of pupae with mites that lay no eggs are the defining elements of VSH. As seen above, these measures are related in colonies with VSH. However, they may not be related when selecting only for hygienic removal of mites. To produce both elements, it may be necessary to base the selective breeding on OVR. Fortunately, measuring the mitevipositing ratio is usually easier than measuring brood removal (one measurement rather than two) and perhaps more accurate (one measurement error rather than two). Unfortunately, mites that lay no eggs and dead mites are easily missed when examining bee pupae.
When measuring OVR (the number of pupae with ovipositing mites / the number of pupae with nonovipositing mites) it is only necessary to determine whether a mite-infested pupa (>7 days postcapping) has any progeny. It is not necessary to count progeny, determine the number of foundress mites, or determine if the progeny could reach adulthood. If a colony does not have enough mites to evaluate for VSH, infested worker brood can be put into a colony, and pupae aged 7 – 10 days postcapping can be evaluated seven days later (see materials and methods).
The presence of VSH is best selected from colonies that have mite ovipositing ratios of 2 or less. Colonies with OVRs > 3 provide unreliable measures of the presence of VSH because natural variation can range from 4 – 10 and perhaps even lower. In this study, the 14 colonies in the H group had mite ovipositing ratios that ranged from 0 to 1.4. The L group ranged from 3.3 to 9.5, and 8 of the 12 colonies showed no measurable expression of VSH in the form of removal of infested brood or a low OVR. It therefore appears that bees with 50 – 100% of the VSH alleles express a significant level of mite resistance, whereas those with 0 – 50% usually do not. This explains why we have produced a significant level of mite resistance in colonies containing VSH queens that were free mated with unselected drones (Harbo and Harris, 2001).
This study did not examine the fate of mites that had their reproductive cycle disrupted. The longevity and subsequent fertility of mites may be affected by this sort of disruption, and in an earlier study we recorded a spike in the frequency of mites that were entrapped between the cell wall and the cocoon (Harris and Harbo, 2000). Ibrahim and Spivak (2006) found that mites taken from bees with VSH (SMR) had a higher frequency of nonoviposition than mites taken from control colonies. Thus the disruption of a mite’s reproductive phase in a brood cell (as produced by VSH) may contribute to reproductive failure or death of the mite if she re-enters a brood cell.
If bees with VSH remove only the pupae from cells infested with mites that lay eggs, then the bees are responding (directly or indirectly) to eggs or ovipositing mites. Indirect responses, for example, could be adult bees detecting chemical or behavioural cues from an ovipositing mite, immature mites or from stressed pupae (Boecking and Spivak, 1999; Boot et al., 1999; Aumeier and Rosenkranz, 2001; Martin et al., 2002; Vandame et al., 2002; Nazzi et al., 2004), or from changes in the cuticular profiles of host pupae (Salvy et al., 2001; Martin et al., 2002).
Harbo, Harris
Acknowledgements
Daniel Winfrey and David Dodge (ARS-USDA, Baton Rouge) provided technical support. Deborah Boykin, statistician, ARS/USDA, Stoneville, MS provided statistical advice. The work was done in cooperation with the Louisiana Agricultural Experiment Station.
References
AUMEIER, P; ROSENKRANZ, P (2001) Scent or movement of Varroa destructor mites does not elicit hygienic behaviour by Africanized and Carniolan honey bees. Apidologie 32: 253-63.
BOECKING, O; SPIVAK, M (1999) Behavioural defences of honey bees against Varroa jacobsoni Oud. Apidologie 30: 141-58.
BOOT, W J; CALIS, J N M; BEETSMA, J; HAI, D M; LAN, N K; TOAN, T V; TRUNG, L Q; MINH, N H (1999) Natural selection of Varroa jacobsoni explains the different reproductive strategies in colonies of Apis cerana and Apis mellifera. Experimental and Applied Acarology 23: 133-44.
CORRÉA-MARQUES, M H; DE JONG, D (1998) Uncapping of worker bee brood, a component of the hygienic behaviour of Africanized honey bees against the mite Varroa jacobsoni Oudemans.
Apidologie 29: 283-90.
ELLIS, J D; DELAPLANE, K S; RICHARDS, C S; HEPBURN, R; BERRY, J A; ELZEN, P J (2004) Hygienic behaviour of Cape and European Apis mellifera (Hymenoptera: Apidae) toward Aethina tumida (Coleoptera: Nitidulidae) eggs oviposited in sealed bee brood.
Annals of the Entomological Society of America 97: 860-864. ELLIS, J D; RICHARDS, C S; HEPBURN, H R; ELZEN, P J (2003)
Oviposition by small hive beetles elicits hygienic responses from Cape honey bees. Naturwissenschaften 90: 532-35.
GILLIAM, M; TABER, S III; RICHARDSON, G V (1983) Hygienic behaviour of honey bees in relation to chalkbrood disease.
Apidologie 14: 29-39.
HARBO, J R; HARRIS, J W (1999) Selecting honey bees for resistance to Varroa jacobsoni. Apidologie 30: 183-196.
HARBO, J R; HARRIS, J W (2001) Resistance to Varroa destructor (Mesostigmata: Varroidae) when mite-resistant queen honey bees (Hymenoptera: Apidae) were free-mated with unselected drones.
Journal of Economic Entomology 94: 1319-1323.
HARBO, J R; HARRIS, J W (2005) Suppressed mite reproduction explained by the behaviour of adult bees. Journal of Apicultural Research 44: 21-23.
HARBO, J R; HARRIS, J W (2005a) The number of genes involved in the SMR trait. American Bee Journal 145: 430.
esponse to Varroa by bees with VSH
HARBO, J R; HOOPINGARNER, R A (1997) Honey bees (Hymenoptera: Apidae) in the United States that express resistance to Varroa jacobsoni (Mesostigmata: Varroidae). Journal of Economic Entomology 90: 893-898.
HARRIS, J W (2008) Effect of brood type on Varroa Sensitive Hygiene by worker honey bees (Hymenoptera: Apidae). Annals of the Entomological Society of America 101: 1137-1144.
HARRIS, J W; HARBO, J R (2000) Changes in reproduction of Varroa destructor after honey bee queens were exchanged between resistant and susceptible colonies. Apidologie 31: 689-699.
IBRAHIM, A; SPIVAK, M (2004) The relationship between suppression of mite reproduction (SMR) and hygienic behaviour. American Bee Journal 144(5): 406.
IBRAHIM, A; SPIVAK, M (2006) The relationship between hygienic behaviour and suppression of mite reproduction as honey bee (Apis mellifera) mechanisms of resistance to Varroa destructor. Apidologie 37: 1-7.
LAPIDGE, K; OLDROYD, B; SPIVAK, M (2002) Seven suggestive quantitative trait loci influence hygienic behaviour in honey bees.
Naturwissenshaften 89: 565-568.
MARTIN, C ; PROVOST, É; BAGNÈRES, A-G ; ROUX, M, CLEMENT, J-L ; LE CONTE, Y (2002) Potential mechanism for detection by Apis mellifera of the parasitic mite Varroa destructor inside sealed brood cells. Physiological Entomology 27: 175-188.
NAZZI, F; VEDOVA, G D; D’AGARO, M (2004) A semiochemical from brood cells infested by Varroa destructor triggers hygienic behaviour in Apis mellifera. Apidologie 35: 65-70.
NEUMANN, P; HÄRTEL, S (2004) Removal of small hive beetle (Aethina tumida) eggs and larvae by African honey bee colonies (Apis mellifera scutellata). Apidologie 35: 31-36.
NEWTON, D C; CANTWELL, G C; BOURQUIN, E P (1975) Removal of freeze-killed brood as an index of nest cleaning behaviour in honey bee colonies (Apis mellifera L). American Bee Journal 115: 388, 402, 406.
161
ROTHENBUHLER, W C (1964) Behaviour genetics of nest cleaning in honey bees. IV. Responses to F1 and backcross generations to disease-killed brood. American Zoologist 4: 111-123.
SALVY, M ; MARTIN, C; BAGNÈRES, A-G; PROVOST, É; ROUX, M ; LE CONTE, Y ; CLEMENT, J L (2001) Modifications of the cuticular hydrocarbon profile of Apis mellifera worker bees in the presence of the ectoparasitic mite Varroa jacobsoni in brood cells.
Parasitology 122: 145-59.
SAS INSTITUTE (2000) OnlineDoc, version 8. SAS Institute Inc., Cary, NC, USA.
SPIVAK, M; DOWNEY, D L (1998) Field assays for hygienic behaviour in honey bees (Hymenoptera: Apidae). Journal of Economic Entomology 91: 64 – 70.
SPIVAK, M; GILLIAM, M (1993) Facultative expression of hygienic behaviour of honey bees in relation to disease resistance. Journal of Apicultural Research 32: 147-157.
SPIVAK, M; REUTER, G S (2001) Resistance to American foulbrood disease by honey bee colonies (Apis mellifera) bred for hygienic behaviour. Apidologie 32: 555-565.
TABER, S. III (1986) Breeding bees resistant to chalkbrood disease.
American Bee Journal 126: 823-825.
VANDAME, R; MORAND, S; COLIN, M E; BELZUNCES, L P (2002) Parasitism in the social bee Apis mellifera: quantifying costs and benefits of behavioural resistance to Varroa destructor mites.
Apidologie 33: 433-45.
VILLEGAS, A J; VILLA, J D (2006) Uncapping of pupal cells by European bees in the United States as responses to Varroa destructor and Galleria mellonella. Journal of Apicultural Research
45: 203-6.
WOODROW, A W; HOLST, E C (1942) The mechanism of colony resistance to American foulbrood. Journal of Economic Entomology 35: 327-330.